结构生物学|折叠体,向超越生物学进军
许多人认为生命的奥秘在于那些折叠的分子 。 现在 , 化学家正向天然蛋白质学习 , 力图人工生成拥有巨大应用潜力的“折叠体” , 但他们能成功吗?科学作家拉谢尔·布拉西尔采访了几位超分子化学家 , 看看他们在忙活什么 。
撰文 | 拉谢尔·布拉西尔(Rachel Brazil)
编译 | 顾淼飞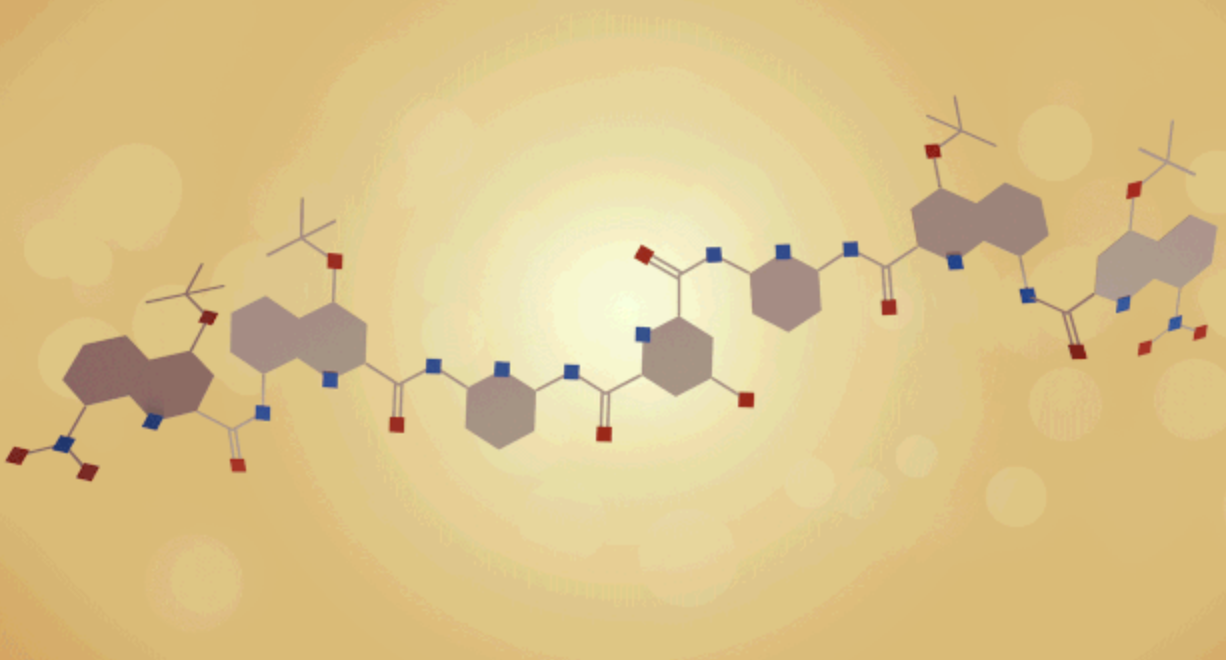
折叠的分子 , 构成了生命的基础 。 用美国威斯康辛大学麦迪孙分校的萨姆·格尔曼(Sam Gellman)的话说:“如果你从化学的角度看待生物学 , 你就无法否认 , 生物学在分子水平上做的几乎每一件复杂的事 , 都是借由序列特异的、折叠的杂聚物来实现的 。 ” 。
如今 , 属于生物学的分子折叠技巧已经被化学家学了来 。 不过 , 英国布里斯托大学的乔纳森·克莱登(Jonathan Clayden)曾经“撂下狠话” , 化学家其实有着更大的野心 , 他们的目的不只是复制出那些天然就存在于生命体中的聚合物 , 还要通过独出心裁的化学设计 , 得到比天然分子更多样、更智能、更“好”的折叠分子 。 你可以说这是青出于蓝而胜于蓝 , 也可以说 , 这是想在对手最擅长的赛道上跑赢对手 。
有望让化学家实现这一野心的 , 是一种叫作“折叠体”的分子(foldamer) , 它也许有朝一日能在这场折叠竞赛中战胜生物学 。
在蛋白质后面跟跑 , 跟着跟着就被落下了
想要合成折叠分子的这个念头 , 最早应该来自于20世纪80年代关于蛋白质折叠的工作 。 格尔曼是最早的实践者之一 , 也是第一位提出“折叠体”一词的人 。 他认为 , “其中的关键性贡献之一 , 是蛋白质建模专家肯·迪尔(Ken Dill)的成果 。 ”目前在纽约州立大学石溪分校的迪尔 , 一直致力于蛋白质折叠的研究 。 他认为 , 蛋白质的折叠过程是因为氨基酸同时具有疏水基和亲水基 。 此前 , 人们一直认为是氢键支配着蛋白质如何形成自身结构 。 然而 , 他的同事罗恩·楚克曼(Ron Zuckermann)用实验证明了事实并非如此 。 楚克曼的反例是由聚-N-取代甘氨酸组成的类肽 , 在它的分子结构上 , 侧链连接在主链上的氮原子上 , 而不是碳原子上 , 它在没有氢键的情况下依然形成了稳定的螺旋状结构 。 这一点让迪尔和楚克曼确信 , 折叠主要是由氨基酸侧链的固有特性造就的 , 氢键提供的结合力充其量只是个聊胜于无的“胶水” 。
除了这个例子 , 还有哪些分子能像类肽那样折叠起来呢?格尔曼想知道这个问题的答案 。 他曾经在一次会议报告之后问过迪尔:“如果我用疏水的苯乙烯和亲水的苯乙烯得到一个聚苯乙烯 , 那么它能折叠吗?”迪尔给出的答案是——是的 , 我认为是可以的 。
在迪尔和楚克曼看来 , 生命始于折叠 , 一个个化学分子能组成生命体 , 其原因也是折叠 。 尽管主流观点都把RNA的复制看作生命过程中的第一步 , 认为正是由于RNA的自主复制 , 才启动了诸如DNA复制、蛋白质合成等一系列分子折叠的进程 。 但是迪尔却认为 , 一定另有一个分子折叠的阶段发生在RNA复制之前 。 他于2017年发表了折叠体假说[1] 。 在他的研究工作中 , 单体被分为两类 , 一类是具有亲水侧链的 , 另一类是具有疏水侧链的 。 他用一个简单的计算模型创建了同时具有这两类单体的化学链 , 然后发现 , 即使是一条很短的链 , 也可以折叠成更紧凑的结构 。
迪尔对此给出了进一步的解释 , 这是因为折叠后的结构使得一种类似于位点的东西——迪尔管它叫“着陆架”(landing pad)——暴露了出来 , 而这些着陆架可以催化生成其他聚合物 , 于是这个过程就有点像多米诺骨牌似的自发延续了下去 , 最终的结果就是产生了原始酶 。 说得再通俗一些 , 如果我们把反应底物比作一锅汤 , 那么在这锅汤里 , 由于疏水单体和亲水单体的序列足够多种多样 , 最后将自发出现全部可能的酶 。 当然 , 从生物学的角度来看 , 生命要以信息存储在DNA上才能开始起算 , 其实在那之前 , 折叠已经发生了 。
所以 , 如果生物学是擅长折叠的化学大师 , 那么真·化学大师可以玩转这个技巧吗?
格尔曼在上世纪90年代开始尝试着合成能折叠的分子 , 并且提出了“折叠体”这个名字用来称呼这类分子 。 但是他随后就发现 , 最开始被看好的聚苯乙烯并不能扛起大旗——用聚苯乙烯来合成这样的折叠分子是行不通的 。 “因为没有人能控制聚苯乙烯中哪个单体会出现在哪里 , 也不知道该怎么控制 , 所以我们后来把研究的焦点放在了聚酰胺身上 。 ”格尔曼解释道 。 他把目光放在了β-氨基酸 , 虽然它的氨基基团不像天然氨基酸那样连接到α-碳上 , 而是连接到β碳上 , 但是由它折叠成的螺旋状分子完全可以跟蛋白质的折叠结构相媲美 。
德国慕尼黑大学的超分子化学家伊万·于克(Ivan Huc)等人也设计出了更多不走寻常路的折叠结构 。 他们的原料一是芳香族的寡聚酰胺;二是具有蛋白原侧链的单体 , 因其有折叠潜力(或者说具有折叠倾向) 。 于克得到的分子是螺旋状的 , 我们可以把它的样子想象成一圈一气呵成削下来、完全没有断掉的苹果皮 。 这圈“苹果皮”的直径可以根据单体尺寸进行调整 , “苹果皮”中间空出来的那个“洞”里 , 可以安置一个“做客”的分子[2] 。分页标题
然而 , 如今折叠体的设计仍然发展得磕磕绊绊 。 尽管计算工具不断发展 , 但还是不如蛋白质和肽的建模工具那么先进 。 “我们一直跟在蛋白质后面跑 , 但是现在被落下了有点远 。 ”于克说 。
催化和制药 , 两手都要抓
格尔曼曾说:“我们最想实现的梦想之一 , 就是创造具有高效催化活性的折叠体 。 ”他最近接下了这项艰巨的挑战 。
在某些情形下 , 酶——这种具有三级结构的分子——能让反应速率加快100万倍 。 尽管格尔曼还不能制备出像酶一样的三级结构 , 但是他确实创造了一种折叠体——能让两个官能团相邻地排列起来 , 从而形成螺旋状的结构[3] 。 格尔曼的折叠体中含有α和β氨基酸 , 包括具有五元环的β残基 , β残基的作用是约束主链的灵活性 , 从而使折叠体的螺旋结构趋于稳定 。
这种折叠体被用来催化生成15个原子以上的大环 , 这类大环分子可用于制药 , 应用潜力巨大 , 可是它们很难制备 , 因为长链分子的两端需要靠得非常近才能发生反应 。 有了各自都连接到β残基上的伯胺和仲胺 , 折叠体就能够正确地定位在化学链的两端 , 从而通过羟醛缩合生成碳碳键 , 得到12~22碳环 。 先前的工作已经表明 , 类似的折叠体体系作催化剂时 , 反应速率比起使用小分子催化剂快出了100倍 。 虽然效果立竿见影 , 但是跟自然界最优秀的催化剂——酶相比 , 折叠体的表现仍然差点意思 。
格尔曼等人也致力于研究折叠体如何以药物分子的身份表现更胜生物学一筹 。 “很多肽类也会被用作药物 , 但是它们在体内很快就分解了 。 ”瑞士ImmuPharma制药公司的总裁迪米特里·迪米特里乌(Dimitri Dimitriou)说 , “如果能高效地制备出在结构上类似于肽 , 在性质上却比肽稳定的折叠体 , 那么它在制药方面的潜力将毫不逊于单克隆抗体产业——站在商业立场来看 , 这是很激动人心的 。 ”他对此非常有信心 , 认为折叠体药物在5年内就可以问世 。
格尔曼在2010年联合创立了Longevity Biotech公司 , 研发含有β-氨基酸的肽类药物[4] 。 “这些肽类分子上的β残基只占1/4~1/3 , 但由于这些β残基是沿着主链分布的 , 所以对水解蛋白酶来说 , 剪断它们也是非常费力且进度缓慢的事情 。 ”他解释道 。 格尔曼的公司管这种折叠体叫作“混血肽(hybridtides)” 。 混血肽的“1.0版” , 是一种能够连接到G蛋白偶联受体的药物 , G蛋白偶联受体是一种跨膜蛋白 , 当受到外部分子的刺激时能在细胞内部传输信号 。 他们目前正进行一项帕金森症候选药物的临床前生物标志物研究 。
在新冠疫情期间 , 格尔曼似乎找到了新的研究方向 , 他开始着手研究能阻断新冠病毒的折叠体药物 , 灵感来自于2009年一项关于艾滋病药物恩夫韦肽的工作[5] 。 恩夫韦肽有36个残基 , 可以高效地阻断病毒吸附到细胞上的这个过程 。 但是这个药物的半衰期实在太短了 , 患者必须每天注射两次 。 格尔曼表示 , “因为α-β主链的因素 , 变体受水解蛋白酶的影响很小 , 影响程度降低到了1/300 , 我们打算用同样的办法来对付新冠病毒 。 ”
跟Longevity Biotech同期开展折叠体药物研究的 , 还有ImmuPharma及其子公司Ureka 。 跟格尔曼的做法不同的是 , 他们的折叠体将部分氨基酸取代成了尿素 。 Ureka的研发总监塞巴斯蒂安·古德罗(Sebastien Goudreau)解释道:“寡聚脲很适合用来生成螺旋结构 , 而且 , 由寡聚脲生成的螺旋结构跟肽的结构很相似的……得到的这个折叠体简直绝了:它既有硬度——这是由尿素主链贡献的 , 也有相当的灵活度——这是由侧链基团贡献的 , 而后者可以像氨基酸那样被取代 。 ”
处于概念验证阶段的Ureka选择了从胰高血糖素样肽-1(GLP-1)起步——这种激素能促进胰岛素的分泌 , 并用来治疗2型糖尿病和非酒精性脂肪肝 。 Ureka的折叠体是用3个尿素残基取代了GLP-1中的连续4个氨基酸[6] 。 迪米特里乌表示:“我们已经证明这是奏效的 , 也证明了这在小鼠体内能大幅延长半衰期 。 ”半衰期的延长意味着药物剂量的减少 , 而且理论上来说 , 如果这种药物能抵抗酶的消化作用的话 , 它甚至可以口服 。
会动的折叠体
自然不仅仅缔造了折叠体 , 还创造了那些能改变自身形状的分子 。 例如 , G蛋白偶联受体(GPCR)在响应激素或是其他能够刺激味觉、嗅觉的分子时 , 可以进行构象转换 。 克莱登已经试着用折叠体来重现这种行为 , 他说:“我们已经设计出具有这样特征的分子——当它们与一个配体结合之后 , 形状会发生改变 , 而分子结构的改变可以用来传递信息……这就是我们所说的动态折叠体(dynamic foldamer) 。 ”
克莱登的研究始于一种非手性的氨基酸——α-氨基异丁酸(AIB) 。 他说:“合成得到的螺旋结构肯定要么是左旋的 , 要么是右旋的 , 事实上它能非常迅速地在这两种结构之间互相转换 。 ”之所以能转换 , 是因为折叠体的胺基端有一个庞大的、呈周期出现的氨基硼酸基团 , 如果与一个同样庞大的手性二醇配体结合 , 它将生成硼酸盐酯 , 而硼酸盐酯将进一步生成连接胺基团的甲醇桥 , 配体的空间位阻驱使折叠体转换到另一种构象[7] 。分页标题
动态折叠体的这种性质有个绝佳的妙用 , 就是用作“智能”药物 , 因为它能根据特定的刺激源在细胞内部通过左旋或右旋来实现“开”或“关”的操作(例如用左旋表示“开” , 用右旋表示“关”) , 从而控制酶的通路 。 克莱登已经证明 , 动态折叠体作为人工受体嵌入磷脂囊泡时是能够正常工作的[8]——这至少说明该思路是可行的 。 他希望未来能将这个概念用到真正的细胞中 。
采用同样的方法 , 克莱登还模拟了我们的色觉 。 在人体中 , 色觉的产生依靠视杆视紫红质上的GPCR受体 , 而克莱登用来替代GPCR受体的分子是偶氮苯生色团 , 它连在AIB折叠体上 , 能在偶氮苯响应光线时改变形状——例如 , 在紫外线条件下 , 这个分子就可以转换到顺式构象 。 克莱登将这一过程形象地称为“构象光电二极管”[9] 。 而这还不是动态折叠体最奇妙的应用 , 更进一步地 , 动态折叠体可以构建“智能”的化学体系 。 例如 , 我们可以用不同颜色的光线来触发反应或终止反应 , 或者控制产物的构象——如果不满意 , 就让它转换成相应的对映体好了 。 又如 , 克莱登正在尝试这样一种体系 , 它能与某个催化剂相结合 , 但是一旦转换构象之后又能把这个催化剂释放出来 , 这一概念可以用于释放类似于酶类抑制剂的物质 。
向三级结构进发
既然折叠体的“竞争对手”是天然的蛋白质 , 那么折叠体的目标 , 自然也就是向着蛋白质所拥有的三级结构进发 。 但是复制类似于蛋白质的三级结构是相当困难的 。 难点之一在于 , 大多数蛋白质有100多个残基 , 这对于化学合成来说几乎是不可能完成的任务;难点之二 , 是折叠体所需要的单体也常常很难合成 。
尽管困难重重 , 化学家还是尝试了一些简单的三级结构 。 他们已经利用一些基团生成了了能模仿锌指结构域(能协调一个或多个锌离子并结合多种生物分子的蛋白质基元)的折叠体;于克甚至在非极性溶剂中生成了螺旋束(helical bundles) , 表明这种结构在与自然界截然不同的环境中也能形成[10] 。
为了创造更大更复杂的结构 , 于克提出要向自然界取经 , 而他选择的“师父”是核糖体 。 核糖体堪称细胞里的蛋白质工厂 , 它里面的整套工作流程涉及mRNA和tRNA的互补配对、氨基酸转运、多肽合成等多个过程 , 有很多值得借鉴的地方 。 于克形容他的长远目标是“操纵核糖体这台机器 , 让它去生产化学物质” 。
于克从2018年开始了关于核糖体的工作 , 他使用大肠杆菌的核糖体合成了一个折叠体和肽的杂化物 , 并且或多或少实现了他的一些设想[11] 。 当然 , 于克的蓝图现在还没有完成 , 未来也不见得容易 。
去做一些自然界力所不能及的事情
退回到30年前 , 一提到分子的折叠 , 人们想到的问题大概还是“生物学上的杂聚物及其折叠能力是否是独一无二的” 。 如今 , 化学家已经给出了答案 , 他们可以举出很多例子 , 来证明化学合成的分子链也具有折叠能力 , 于是 , 问题变成了“能否制备足够复杂的大型折叠分子 , 以及能否操纵它们” 。
折叠体研究领域的进步当然得益于向自然界的“偷师学艺” , 但是化学家确实从中找到了更广阔的空间 。 “我们需要思考一些自然界根本就办不到的事情 。 ”于克提出 , “至于关键性的进展将会出现在哪里 , 谁知道呢?”
参考文献
[1] E Guseva, R N Zuckermann and K A Dill, Proc. Natl Acad. Sci. USA, 2017, 114, E7460 (DOI: 10.1073/pnas.1620179114 )
[2] J Garric, J-M Léger and I Huc, Angew. Chem. Int. Ed., 2005, 44, 1954 (DOI:10.1002/anie.200462898 )
[3] Z C Girvin, M K Andrews, X Liu, S H Gellman, Science, 2019, 366, 1528 (DOI:10.1126/science.aax7344 )
【结构生物学|折叠体,向超越生物学进军】[4] R Cheloha et al, Nat. Biotechnol., 2014, 32, 653 (DOI: 10.1038/nbt.2920 )
[5] S W Horne et al, Proc. Natl Acad. Sci. USA, 2009, 106, 14751 (DOI:10.1073/pnas.0902663106 )
[6] J Fremaux et al, Nat Commun., 2019, 10, 924 (DOI: 10.1038/s41467-019-08793-y )
[7] R Brown et al, Nat. Chem., 2013, 5, 853 (DOI: 10.1038/nchem.1747 )
[8] F Lister et al, Nat. Chem., 2017, 9, 420 (DOI: 10.1038/nchem.2736 )
[9] D Mazzier et al, J. Am. Chem. Soc., 2016, 138, 8007 (DOI:10.1021/jacs.6b04435 )
[10] S De et al, Nat. Chem., 2018, 10, 51 (DOI: 10.1038/nchem.2854 )
[11] J M Rogers et al, Nat. Chem., 2018, 10, 405 (DOI: 10.1038/s41557-018-0007-x )
原文链接:https://www.chemistryworld.com/features/the-function-of-folding/4012081.article
- |中石化又投资一新能源企业!石油企业产业结构正发生转变
- 躺着看电影|山东优化事业单位岗位设置治理 将15个行业结构比例指导尺度整合为6类
- 健识局|次均费用上涨,公立医院收入结构大变:门诊和出院人次双双下降
- 2.5 证明结构力
- FlexPai|国产折叠屏手机为何成功出圈?FlexPai 2价格优势率先成为普及先锋
- 中国汽车报网|奔驰发布全新战略:聚焦全系电动化 强化结构性盈利
- 市场|市场抢夺战迫在眉睫,户外LED广告机渠道结构发生重大变化
- 负债率|稳降负债率,金辉持续优化债务结构
- 鸟类|鸟类的消化系统与结构
- 花生|结构调整促河北枣强农民节水增效