按关键词阅读:
人类生理和行为的昼夜节律是由细胞中时钟蛋白的周期性相互作用而驱动的 。 当时钟蛋白出现突变时 , 昼夜节律可能就会失调 , 有的人往往会比平时更早入睡和起床 , 有的人则会成为典型的夜猫子 , 通常要熬夜到2点之后才能入睡 , 当然也难以早起 。
2017年来自美国洛克菲勒大学的团队发现了一个影响人体日常节奏的生物钟的关键成分的变体:即缺失外显子11的编码隐花色素的蛋白CRY1( CRY1Δ11) , 该蛋白的遗传变异导致了延迟入睡的迟睡眠阶段障碍(DSPD) 。 但其中CRY1的突变是如何影响人类昼夜节律的仍未可知 。
近日 , 来自美国加利福尼亚大学的研究人员在《PNAS》上发表了题为The human CRY1 tail controls circadian timing by regulating its association with CLOCK:BMAL1的研究成果 , 揭示了生物钟蛋白之一CRY1的外显子11缺失引发延迟睡眠相障碍的机制 , 即通过调节CRY1与CLOCK:BMAL1的互作紧密程度而改变了昼夜节律 。

文章图片
https://doi.org/10.1073/pnas.1920653117
前期研究发现 , 控制人体昼夜节律的时钟蛋白主要有4种 , 时钟蛋白CLOCK和BMAL1能够形成异二聚体转录因子 , 促进CRY1/2和PER1/2的表达 , 而CRY1/2和PER1/2又能够结合成复合物来抑制前两个蛋白的活性 , 由此构成了一个调控生物钟的反馈回路 。 而改变CRY1对BMAL1中TAD结构域的亲和力 , 可以通过缩短或延长反馈回路中抑制的持续时间而改变昼夜节律 。
为了明确CRY1的外显子11的突变是如何影响昼夜节律的 , 研究人员首先使用荧光偏振(FP)对CRY1的尾巴与反式PHR功能域的结合进行了分析 , 发现CRY1尾部可以直接与CRY1的PHR功能域结合 , 而外显子11缺失后 , 其与PHR的亲和力下降了4倍 。
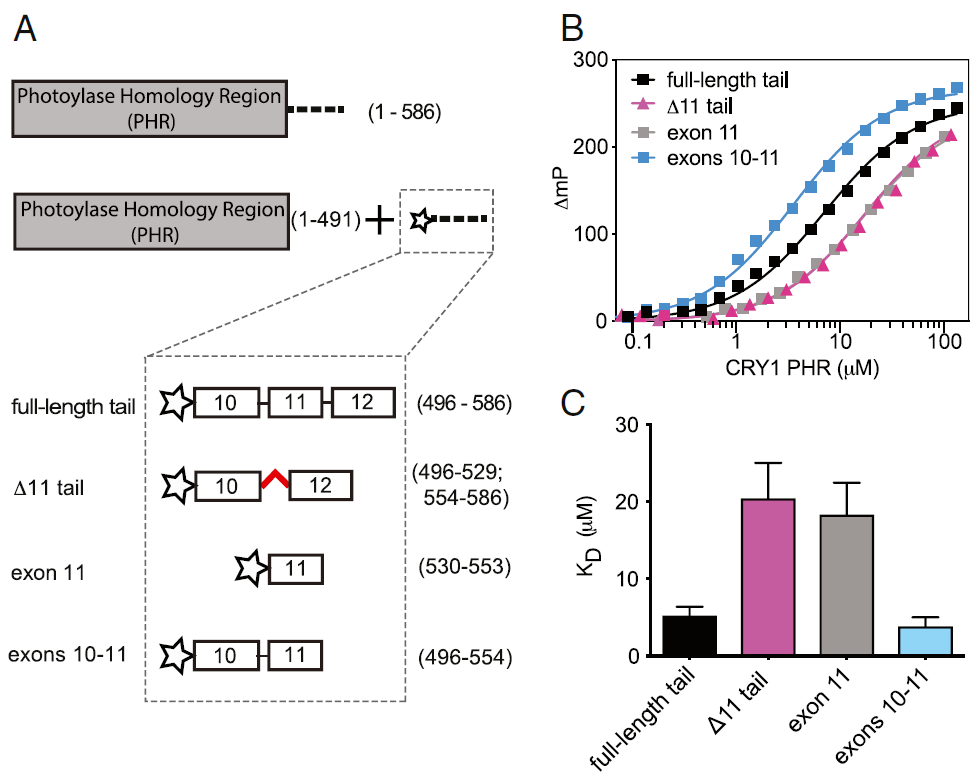
文章图片
CRY1尾部的外显子10和11是PHR结合所必需的
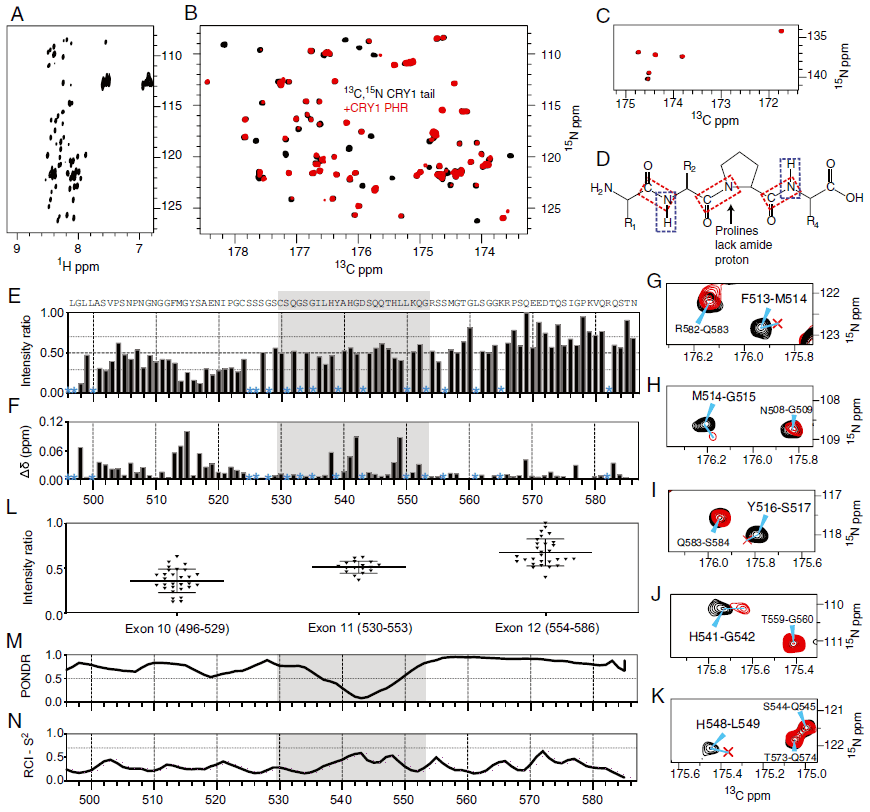
文章图片
核磁共振波谱绘制了CRY1尾部的PHR结合位点
【障碍|PNAS:“夜猫子”睡眠障碍竟然会是基因突变导致的!】CRY1 PHR与CLOCK:BMAL1异源二聚体的结合有两个接口 , 一个是CRY1的卷曲螺旋与BMAL1的TAD(反式激活功能域)结合 , 另一个是CRY1与CLOCK的PAS-B结合 。 基于外显子11在CRY1尾巴与其PHR结构域互作中的重要作用 , 研究人员使用荧光标记的BMAL1 TAD和CLOCK PAS-B进行了基于FP的PHR结合力分析 , 发现CRY1尾部在调节CRY1 PHR和BMAL1 TAD互作中无明显作用 , 但其可以从PHR域中置换出CLOCK PAS-B , 且其中外显子11对调节CLOCK PAS-B和CRY1 PHR结构域之间的互作具有关键重要作用 。
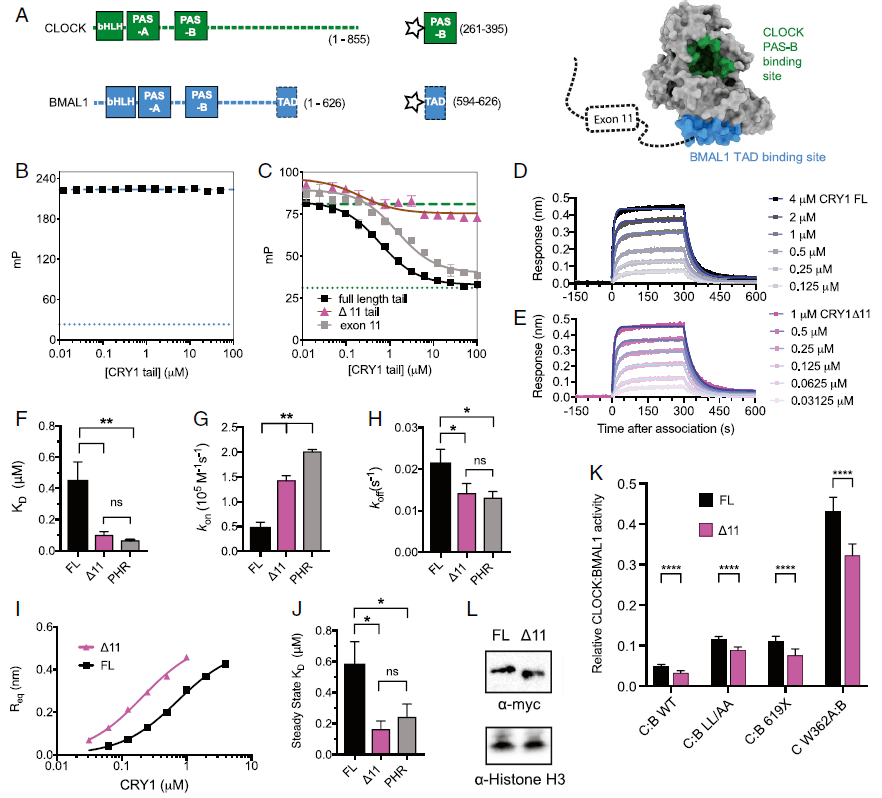
文章图片
外显子11与CLOCK PAS-B竞争 , 降低CRY1与CLOCK:BMAL1的亲和力
之后 , 研究人员使用生物层干涉术(BLI)来定量评估全长CRY1 , CRY1Δ11和无尾CRY1 PHR域对CLOCK:BMAL1的PAS核心域的亲和力 , 发现与全长CRY1相比 , CRY1Δ11和无尾CRY1 PHR域与其具有更高的亲和力 。 并通过荧光素酶报告系统证明了CRY1Δ11对CLOCK的亲和力增强直接增强了其抑制CLOCK:BMAL1转录激活的能力 。 由此 , 拉长了调控生物钟的反馈回路中抑制的持续时间 , 进而改变了昼夜节律 , 导致延迟入睡 。
该研究的通讯作者Carrie L. Partch表示:“研究结果表明CRY1与CLOCK互作的紧密程度决定了生物时钟运行的速度 , 这也在提示我们 , 应该寻找能够与CLOCK:BMAL1结合并可以起到与CRY1尾巴相同功能的药物 , 开发出可以缩短睡眠阶段障碍患者时钟的疗法 。 ”![]()
稿源:(健康界)
【】网址:http://www.shadafang.com/c/hn1104c4Z62020.html
标题:障碍|PNAS:“夜猫子”睡眠障碍竟然会是基因突变导致的!
