文章图片

文章图片

分类包括了基于知觉特征、语义关系或两者结合相关的多个刺激 。
习得类别表征有助于帮助动物和人类从已经获得的知识中进行推断从而对新奇体验做出反应 。 对类别的学习和回忆激活了大量脑区 , 包括感觉皮层 , 这突出了其联想性质 。 联想记忆存储在跨越脑区的分布式网络中 。 然而 , 神经元类别表征的形成是在所有激活的脑区中发生 , 还是仅存储在高级皮质关联区域的子集中尚未清楚 。 近日 , Goltstein研究团队在Nature Neuroscience杂志发表了一项研究成果 , 他们详细描述了大群神经元在整个类别学习过程中对小鼠视觉皮层九个区域的反应特性 , 并发现学习不仅会导致新生神经元对选择和奖励的反应 , 还会导致刺激和类别调整的变化 , 从而增强对所学视觉类别的辨别能力 。
小鼠辨别、概括和记忆视觉类别
为了测试小鼠学习视觉类别的能力 , 研究人员在触摸屏操作室中训练了8只雄性小鼠 , 以区分一组方向和空间频率不同的42种光栅刺激 。 通过对角类别边界将二维刺激空间划分成奖励和非奖励类别 。 这种信息整合分类的特征是需要同时权衡多个刺激特征维度 。 根据设计 , 分类任务具有知觉成分(辨别方向和空间频率)和语义成分(多个刺激共享相同的意义) 。 动物被训练了4到6天的时间来区分两个离类别边界最远的刺激 。 一旦小鼠辨别出这些初始刺激的概率很高 , 就再引入额外的刺激 , 逐渐接近类别边界 , 直到属于两个类别的所有刺激都出现 。 尽管在仅6至8天的时间内添加了40个新刺激 , 但所有小鼠皆可跨越类别边界区分刺激 , 在类别内概括刺激 , 并将其扩展到以前未遇到过的刺激 。 小鼠已经形成了语义记忆 。 Fig1小鼠在触摸屏操作室学习区分信息整合类别
习得的视觉分类部分依赖于视觉区域的可塑性
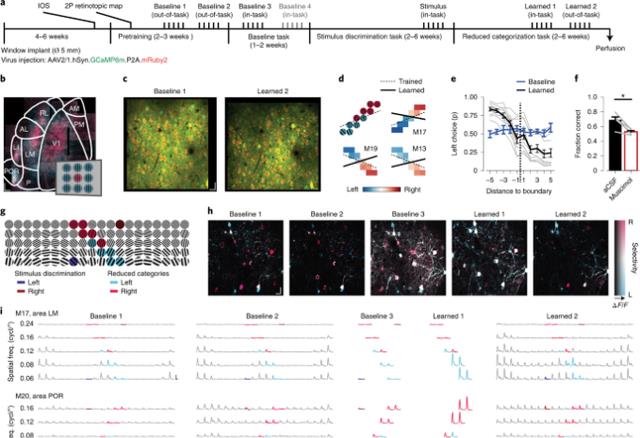
【神经元|联想记忆塑造全脑分布网络的实验证据——基于小鼠视觉皮层研究】接下来 , 研究人员实施了头部固定版本的分类任务 , 提供了对视觉刺激的精确控制 , 并进行双光子检查 。 实验结果表明 , 虽然习得的分类行为并不严格限制在刺激的确切视野位置 , 但它会因改变刺激位置而受损 。 这表明视觉区域至少存储了一定数量的关于所学类别的知觉或语义信息 。 Fig2 头部固定类别学习依赖于早期视觉区域的可塑性
视觉分类取决于视觉皮层的活动
为了详细评估这些区域的神经反应如何随着类别学习而改变 , 研究人员使用长时体内双光子钙成像 , 在整个学习过程中 , 对相同的神经元重复记录 。
Fig3 在整个分类学习过程中 , 多个视觉区域的慢性钙成像
为了准确描述视觉皮层区域的神经元反应在类别学习时的变化 , 研究人员先说明了激活的神经元的总数 。 其次 , 又描述了由这种激活的神经元编码的信息类型 。 发现类别学习后 , 刺激激活神经元和任务激活神经元的比例发生了变化 。
Fig4 类别学习后 , 刺激激活神经元和任务激活神经元的比例发生了变化
新出现的反应神经元是因何而编码的呢?研究人员认为小鼠视皮层的功能比传统假设更加多样化;它可以受到许多视觉刺激以外因素的驱动和调节 。
研究人员使用了一个广义线性模型来估计刺激方向、空间、频率和类别、运动和奖励等对单个神经元放电活动的贡献 。 发现“获得的”神经元 , 除了受到视觉成分的调节外 , 还受到行为选择及奖励的调节 。
Fig5 使用广义线性模型识别单个神经元的刺激、选择、奖励和运动相关活动模式
研究人员还观察到 , 学习后语义CTI增加 , 且在V1和POR区最为明显 。 学习后的特征CTI也普遍增加但无个别区域特别突出 。 在类别学习之前的选择-选择性有助于提高POR区的ΔCTI , 从而对类别的语义而非感知进行调整 。
Fig6 choice-selective PoR神经元具有类别选择性
结 论
Goltstein研究团队利用小鼠视觉类别学习范式发现 , 小鼠学会对视觉刺激进行分类 , 区分不同的类别 , 并在不同的类别下进行概括 。 这种分类能力取决于视皮层中神经元的活动 。
对视觉皮层九个区域进行的长期钙成像发现 , 在学习过程中 , 特征调整和类别调整发生了变化 , 其中最显著的是嗅觉后区域 。 这些结果为联想记忆塑造全脑分布网络的观点提供了证据 , 学习在早期阶段形成知觉表征 , 并在下游支持语义内容 。
- 联想|在电脑城几年的工作经历,谈谈对联想电脑的几点认识
- 卫星|记忆中的考古“大家”|“新中国河南考古第一人”——安金槐先生
- 联想|私有化是联想困局的最佳解决方案?
- 联想|那些说我为联想洗地的人,为何自己不先从所在的无核心技术公司辞职?
- 联想|因为司马南,联想口碑崩了
- 贸易战|联想这些年骚操作不断,到底做错了啥?
- 联想|拆解联想电脑,没有任何一个核心零部件是国产的,塑料壳和螺丝是国产的
- 联想|胡大佬评联想事件,看看司马南如何接招?
- 联想|柳传志和杨元庆并不是罪人,网民的反应过激了
- 联想|司马南与联想之争,发言请用事实说话