我这个人有点强迫症:走在路上,如果觉得哪只鞋比另一只更松一点,我都要立刻蹲下调整到松紧相同。其实我们人类都喜欢对称——春联要挂一对,盖房子要东西厢,从半坡文化的陶瓶到巴黎圣母院的正立面,对称之美在人类文化中随处可见。
显而易见,这恐怕是因为我们自己就是对称的。然而事情并没有这么简单:我们“对称了,但又好像没完全对称”——毕竟,几乎所有人的心脏都偏左,肝脏都偏右……那么,动物身体的对称性,究竟是怎么产生的,又有什么意义呢?
翩若游龙
其实,在地球生命的演化史中,身体结构对称性的“设计”是件具有划时代意义的大事。
三维空间中的生物体,可以是辐射对称、两侧对称或者不对称的。如果有一个贯穿身体的轴,沿着轴所在的多个平面可以将生物切成几部分,每一部分都一样,那么这种生物就是辐射对称的,例如多细胞动物最早分化出的海绵和栉水母动物,以及五辐对称的海星等等;如果找不到这样一个轴,而只有沿着一个平面切才能获得镜像对称的两半,那么这种生物就是两侧对称的。显然,包括人类在内的大多数动物采用了两侧对称的“设计方案”。辐射对称和两侧对称的早期演化,与地质环境不无关系。
动物对称方式,依次为球对称、辐射对称(五辐)、双辐射对称、两侧对称。Fig. 1, Hollo ? G. 2015 A new paradigm for animal symmetry. Interface Focus 5: 20150032.
距今大约5.4亿年前,早期的多细胞动物经历了一次快速的辐射演化,产生了众多的门类,这就是著名的寒武纪大爆发。虽然名为“爆发”,这仍是一场跨越四千万年时间的演化历史剧,而开幕的第一场是“田园牧歌”式的埃迪卡拉纪晚期。在此前漫长的三十亿年中,以蓝细菌为主的单细胞生物日积月累,已经将广阔的浅海地区改造成了铺着一层“微生物席”的广袤“草原”。大部分埃迪卡拉动物靠滤食环境中的营养物质生活,互相之间也没有什么竞争,因此接触环境的表面积越大越好。比如著名的狄更逊水母就是铺在海床上的巨大扁盘;查恩盘虫则有“根”有“叶”,更像是直立在海水里的植物。

文章插图
埃迪卡拉纪典型动物群落想象图。(作者:John Sibbick)
而有的动物则采取了不同的策略。2019年,中科院南京古生物研究所的科学家们在《自然》杂志上发表论文,报道了距今5.5亿年前的夷陵虫Yilingia。这种埃迪卡拉生物可不是靠天吃饭的“酒囊饭袋”,它们身体细长分节,两侧对称,很可能是环节动物或者节肢动物的祖先。夷陵虫们身后拖着的轨迹表明它们在微生物席“大草原”上主动游走牧食,很可能还会潜入微生物席之中栖息。

文章插图
夷陵虫身体(蓝色标记)和运动轨迹(白色标记)化石(上),及复原想象图(下)。Fig. 1f, Fig. 3, Chen et al, Nature, 2019.
这样有方向的主动运动,不难想象只有夷陵虫那两侧对称、分节的身体结构才更拿手。在岁月静好“埃迪卡拉花园”里,金伯拉虫和夷陵虫等两侧对称动物担任起了“除草机”。而进入寒武纪后,越来越多的两侧对称动物出现,它们在不断沉积的微生物席里掘穴潜行,上下而求索,不仅获取食物,也防御前辈们没见过的捕食者。这些活动对在沉寂中积攒数十亿年的微生物席产生了翻天覆地的扰动,被地质学家们称为“海洋底质革命”。
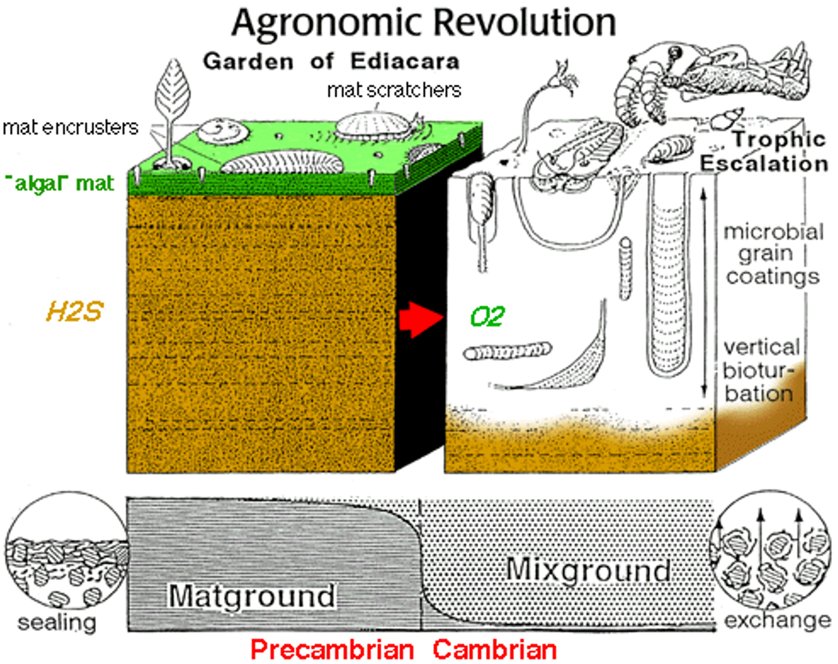
文章插图
底质革命示意图,从埃迪卡拉纪(左)到寒武纪(右)。(图片来源:sepmstrata.org)
有研究提出,在所有对称性设计中,两侧对称能够为动物改变运动方向提供最有效的推动力。可以说,两侧对称的身体构造,为动物们真正“动”起来提供了基础,让地球变成了物种间频繁竞争、捕食的运动场。当然,两侧对称的适应意义不仅限于运动能力,也有研究提出,两侧对称的结构能提高动物体内物质输送的效率。
源远流长
回看历史,我们两侧对称的身体源自多细胞动物的早期演化;而在每一只两侧对称动物的发育过程中,胚胎的对称性也经历了复杂的变化,受到了相关发育基因的精细调控。在发育生物学描述中,两侧对称动物在三维空间中的两个坐标轴上存在差异:区分前端和后端的前后(anterior-posterior,AP)轴,和区分背面与腹面的背腹(dorsal-ventral,DV)轴。体轴的建立与维持,是胚胎发育过程中的大事。
- 韩慧英|见证南北文化交融 山西文水“采茶调”的前世今生
- 周易|让日常生活美起来|隐于市的“江湖人”,午休一小时仗剑走天涯
- 上海|“佳士得上海”迁入新址 外滩再添艺术新空间
- 猪肝|甘甜不上火,护肝又滋补——和“红嘴绿鹦哥”一起迎接初春的暖阳吧!
- 中共一大纪念馆|“伟大建党精神专题展”开启全国巡展!
- 丸子|这是菜市场“最脏”的3种菜,再便宜也不要买,你却天天端上桌
- 艺术培训|德州市文化馆2022年“德艺课堂” 春季公益艺术培训开始招生!
- 文化节|沪上首家传统文化传承中心——“北站传统文化传承中心”揭牌成立
- 艺术|东西方艺术巧妙融合 重庆女孩画“家门皮肤”受年轻人追捧
- 清华大学|美术馆嵌入产业园,上海打开“科创+文创”新模式